Exam Details
Exam Code
:MCAT-TESTExam Name
:Medical College Admission Test: Verbal Reasoning, Biological Sciences, Physical Sciences, Writing SampleCertification
:Medical Tests CertificationsVendor
:Medical TestsTotal Questions
:812 Q&AsLast Updated
:Apr 16, 2025
Medical Tests Medical Tests Certifications MCAT-TEST Questions & Answers
-
Question 611:
Just as the ingestion of nutrients is mandatory for human life, so is the excretion of metabolic waste products. One of these nutrients, protein, is used for building muscle, nucleic acids, and countless compounds integral to homeostasis.
However, the catabolism of the amino acids generated from protein digestion produces ammonia, which, if not further degraded, can become toxic. Similarly, if the same salts that provide energy and chemical balance to cells are in excess,
fluid retention will occur, damaging the circulatory, cardiac, and pulmonary systems.
One of the most important homeostatic organs is the kidney, which closely regulates the excretion and reabsorption of many essential ions and molecules. One mechanism of renal function involves the secretion of antidiuretic hormone
(ADH).
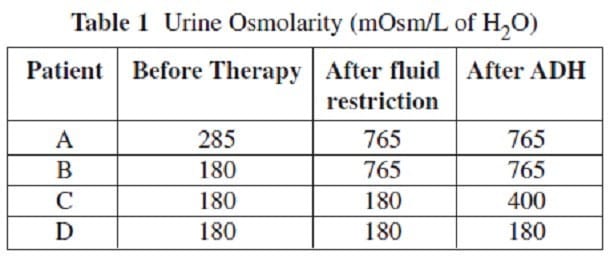
Diabetes insipidus (DI), is the condition that occurs when ADH is ineffective. As a result, the kidneys are unable to concentrate urine, leading to excessive water loss. There are two types of DI -- central and nephrogenic. Central DI occurs when there is a deficiency in the quantity or quality of ADH produced. Nephrogenic DI occurs when the kidney tubules are unresponsive to ADH. To differentiate between these two conditions, a patient's urine osmolarity is measured both prior to therapy and after a 24-hour restriction on fluid intake. Exogenous ADH is then administered and urine osmolarity is measured again. The table below gives the results of testing on four patients. Assume that a urine osmolarity of 285 mOsm/L of H2O is normal.
An elevated and potentially toxic level of ammonia in the blood (hyperammonemia) would most likely result from a defect in an enzyme involved in:
A. glycolysis.
B. fatty acid catabolism.
C. the urea cycle.
D. nucleic acid degradation.
-
Question 612:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.

Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for
than Hb.
In sperm whales, the Mb content of muscle is about 0.004 moles/kg of muscle. If a sperm whale has 1000 kg of muscle, approximately how much O2 is bound to Mb, assuming that the Mb is saturated with O2?
A. 4 moles
B. 8 moles
C. 12 moles
D. 16 moles
-
Question 613:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
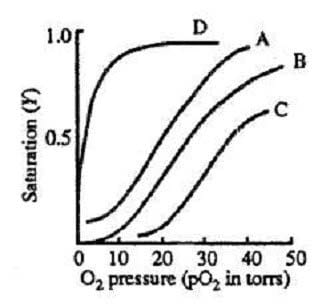
Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
The utilization coefficient is continually being adjusted in response to physiological changes. Which of the following values most likely represents the utilization coefficient for human adult Hb during strenuous exercise?
A. 0.0
B. 0.125
C. 0.25
D. 0.75
-
Question 614:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
Figure 1 Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
A sample of human adult Hb is placed in an 8 M urea solution, resulting in the disruption of noncovalent interactions. After this procedure, the chains of Hb are isolated. Which of the four curves most closely resembles the O2-dissociation curve for the isolated chains? [Note: Assume that Curve B represents the O2-dissociation curve for human adult Hb in vivo.]
A. Curve A
B. Curve B
C. Curve C
D. Curve D
-
Question 615:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
The sigmoidal shape of the O2-dissociation curve of Hb is due to:
A. the effects of oxidation and reduction on the heme groups within the Hb molecule.
B. the concentration of carbon dioxide in the blood.
C. the fact that Hb has a lower affinity for than Mb.
D. the cooperativity in binding among the subunits of the Hb molecule.
-
Question 616:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
If Curve B represents the O2-dissociation curve for human adult Hb, which of the following best explains why Curve A most closely resembles the curve for fetal Hb?
A. Fetal tissue has a higher metabolic rate than adult tissue.
B. Fetal tissue has a lower metabolic rate than adult tissue.
C. Fetal Hb has a higher affinity for than adult Hb.
D. Fetal Hb has a lower affinity for than adult Hb.
-
Question 617:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
If Curve B represents the O2-dissociation curve for elephant Hb, which curve most closely resembles the curve for mouse Hb?
A. Curve A
B. Curve B
C. Curve C
D. Curve D
-
Question 618:
Hemoglobin (Hb) and myoglobin (Mb) are the O2-carrying proteins in vertebrates. Hb, which is contained within red blood cells, serves as the O2 carrier in blood and also plays a vital role in the transport of CO2 and H+. Vertebrate Hb consists of four polypeptides (subunits) each with a heme group. The four chains are held together by noncovalent attractions. The affinity of Hb for O2 varies between species and within species depending on such factors as blood pH, stage of development, and body size. For example, small mammals give up O2 more readily than large mammals because small mammals have a higher metabolic rate and require more O2 per gram of tissue.
The binding of O2 to Hb is also dependent on the cooperativity of the Hb subunits. That is, binding at one heme facilitates the binding of O2 at the other hemes within the Hb molecule by altering the conformation of the entire molecule. This conformational change makes subsequent binding of O2 more energetically favorable. Conversely, the unloading of O2 at one heme facilitates the unloading of O2 at the others by a similar mechanism.
Figure 1 depicts the O2-dissociation curves of Hb (Curves A, B, and C) and myoglobin (Curve D), where saturation, Y, is the fractional occupancy of the O2-binding sites. The fraction of O2 that is transferred from Hb as the blood passes through the tissue capillaries is called the utilization coefficient. A normal value is approximately 0.25.
Figure 1
Myoglobin facilitates transport in muscle and serves as a reserve store of O2. Mb is a single polypeptide chain containing a heme group, with a molecular weight of 18 kd. As can be seen in Figure 1, Mb (Curve D) has a greater affinity for than Hb.
The llama is a warm-blooded mammal that lives in regions of unusually high altitudes, and has evolved a type of Hb that adapts it to such an existence. If Curve B represents the O2-dissociation curve for horse Hb, which curve would most closely resemble the curve for llama Hb?
A. Curve A
B. Curve B
C. Curve C
D. Curve D
-
Question 619:
Saul Hoffman's scientific journal paper published in 2015 in Societies explores the relationship between two topics that at the surface are very distant from each other. As he goes on to state, "It is relatively easy, at least for an economist, to see why economists would be attracted to issues like teen pregnancy and teen childbearing, despite their apparent distance from the core topics of economics. First, economics ?especially microeconomics ?is fundamentally the study of choices that individuals make, traditionally and most often in formal markets with monetary prices, but now more and more frequently outside that sphere. Viewed from that perspective, choices involving sexual and fertility behavior among teens are an incredibly challenging, but inviting, target. Is it possible to identify the role of economic incentives, including government policy, on these behaviors? Is it sensible to apply traditional models of rational choice decision-making to teens?
Second, the traditional concern about teen fertility was predicated on the notion that it was an economically catastrophic act. In a famous and oft-quoted 1968 article, Arthur Campbell wrote that 'The girl who has an illegitimate child at the age of 16 suddenly has 90 percent of her life's script written for her,' including reduced opportunities for schooling, the labor market, and marriage. But it doesn't take too much reflection to appreciate that more may be going on in leading to these poor outcomes than just a teen birth. Disentangling the causal effect of teen childbearing on subsequent socio-economic outcomes from its correlational effect is another deliciously inviting and challenging target, this time well-suited for the applied economist or econometrician.
Just to make all this yet more inviting, the two research strands are closely related. Suppose it could be demonstrated that for some teens the socio-economic impact of a teen birth was negligible. For example, maybe future prospects for some teens were equally poor with or without a birth or perhaps government programs provided substantial benefits, so that the net impact on socio-economic well-being was consequently small or even positive. Then, it might well be 'rational' in an economic sense to have a teen birth in the first place, thereby linking the research on the causal impact of a teen birth with the research on the choice determinants of a teen birth. So what came to be known as the teen birth `causes' literature and the teen birth `consequences' literature were clearly interrelated.
And then, to add yet another layer of challenge, the teen fertility rate in the U.S. has fallen at a rate that is totally unprecedented. Teen fertility was once widespread, with most of it occurring within early and sometimes not entirely voluntary marriage. In 1960, the teen fertility rate was approximately 90 births per 1000, which implied that more than 40% of women ever had a teen birth. When I published my first article on teen births 25 years ago, the teen fertility rate was 60 births per 1000, down one-third from 1960, but it had increased six years in a row in what turned out to be a deviation from the downward trend. Since then the rate has declined every single year, except for a short but puzzling uptick between 2005 and 2007. In 2014, the teen fertility rate was 24.2 births per 1000, the lowest teen fertility rate ever recorded in the U.S., though still shockingly high by European standards. Thus, the rate fell by more than 50% during my professional association with the topic and by 70% since 1960. Of course, at the same time teen marital births largely disappeared, falling from 85% of teen births to 12%.
This adds yet another focus for economic research. Why did the rate fall? Did it have anything to do with changes in the costs of teen childbearing or changes in policy? Is it a good thing or not?
In this article I try to make sense out of these various research strands by providing a personal narrative through the economics literature on teen childbearing, with a special emphasis on the three issues discussed above. My goal is to make the literature, including some reasonably technical content, accessible and valuable to a non-economist."
Hoffman, S. (2015). Teen Childbearing and Economics: A Short History of a 25-Year Research Love Affair. Societies, 5(3), 646-663. doi:10.3390/soc5030646
What is the most likely reason for the author's use of quotation marks around "rational" in paragraph 3?
A. The author disagrees with the idea that teen birth is rational and wishes to distance himself from it.
B. The author disagrees with the way the term "rational" is used to indicate that economists might perceive teen birth as financially beneficial.
C. The possible benefits of a teen birth are so small that it is not necessarily meaningful to describe them as "rational."
D. The choice of having a teen birth might not have been thought out methodically, but makes sense according to a monetary cost/benefit analysis.
-
Question 620:
Saul Hoffman's scientific journal paper published in 2015 in Societies explores the relationship between two topics that at the surface are very distant from each other. As he goes on to state, "It is relatively easy, at least for an economist, to see why economists would be attracted to issues like teen pregnancy and teen childbearing, despite their apparent distance from the core topics of economics. First, economics ?especially microeconomics ?is fundamentally the study of choices that individuals make, traditionally and most often in formal markets with monetary prices, but now more and more frequently outside that sphere. Viewed from that perspective, choices involving sexual and fertility behavior among teens are an incredibly challenging, but inviting, target. Is it possible to identify the role of economic incentives, including government policy, on these behaviors? Is it sensible to apply traditional models of rational choice decision-making to teens?
Second, the traditional concern about teen fertility was predicated on the notion that it was an economically catastrophic act. In a famous and oft-quoted 1968 article, Arthur Campbell wrote that 'The girl who has an illegitimate child at the age of 16 suddenly has 90 percent of her life's script written for her,' including reduced opportunities for schooling, the labor market, and marriage. But it doesn't take too much reflection to appreciate that more may be going on in leading to these poor outcomes than just a teen birth. Disentangling the causal effect of teen childbearing on subsequent socio-economic outcomes from its correlational effect is another deliciously inviting and challenging target, this time well-suited for the applied economist or econometrician.
Just to make all this yet more inviting, the two research strands are closely related. Suppose it could be demonstrated that for some teens the socio-economic impact of a teen birth was negligible. For example, maybe future prospects for some teens were equally poor with or without a birth or perhaps government programs provided substantial benefits, so that the net impact on socio-economic well-being was consequently small or even positive. Then, it might well be 'rational' in an economic sense to have a teen birth in the first place, thereby linking the research on the causal impact of a teen birth with the research on the choice determinants of a teen birth. So what came to be known as the teen birth `causes' literature and the teen birth `consequences' literature were clearly interrelated.
And then, to add yet another layer of challenge, the teen fertility rate in the U.S. has fallen at a rate that is totally unprecedented. Teen fertility was once widespread, with most of it occurring within early and sometimes not entirely voluntary marriage. In 1960, the teen fertility rate was approximately 90 births per 1000, which implied that more than 40% of women ever had a teen birth. When I published my first article on teen births 25 years ago, the teen fertility rate was 60 births per 1000, down one-third from 1960, but it had increased six years in a row in what turned out to be a deviation from the downward trend. Since then the rate has declined every single year, except for a short but puzzling uptick between 2005 and 2007. In 2014, the teen fertility rate was 24.2 births per 1000, the lowest teen fertility rate ever recorded in the U.S., though still shockingly high by European standards. Thus, the rate fell by more than 50% during my professional association with the topic and by 70% since 1960. Of course, at the same time teen marital births largely disappeared, falling from 85% of teen births to 12%.
This adds yet another focus for economic research. Why did the rate fall? Did it have anything to do with changes in the costs of teen childbearing or changes in policy? Is it a good thing or not?
In this article I try to make sense out of these various research strands by providing a personal narrative through the economics literature on teen childbearing, with a special emphasis on the three issues discussed above. My goal is to make the literature, including some reasonably technical content, accessible and valuable to a non-economist."
Hoffman, S. (2015). Teen Childbearing and Economics: A Short History of a 25-Year Research Love Affair. Societies, 5(3), 646-663. doi:10.3390/soc5030646
The author would likely view the choice of whether to use birth control as:
A. a personal choice that should be governed by one's beliefs about the morality of using it.
B. a decision governed by cost-benefit analyses including factors such as cost of the birth control.
C. a sign of innate intelligence.
D. an ethical choice involving having children only when one can provide them with a good life.
Related Exams:
Tips on How to Prepare for the Exams
Nowadays, the certification exams become more and more important and required by more and more enterprises when applying for a job. But how to prepare for the exam effectively? How to prepare for the exam in a short time with less efforts? How to get a ideal result and how to find the most reliable resources? Here on Vcedump.com, you will find all the answers. Vcedump.com provide not only Medical Tests exam questions, answers and explanations but also complete assistance on your exam preparation and certification application. If you are confused on your MCAT-TEST exam preparations and Medical Tests certification application, do not hesitate to visit our Vcedump.com to find your solutions here.